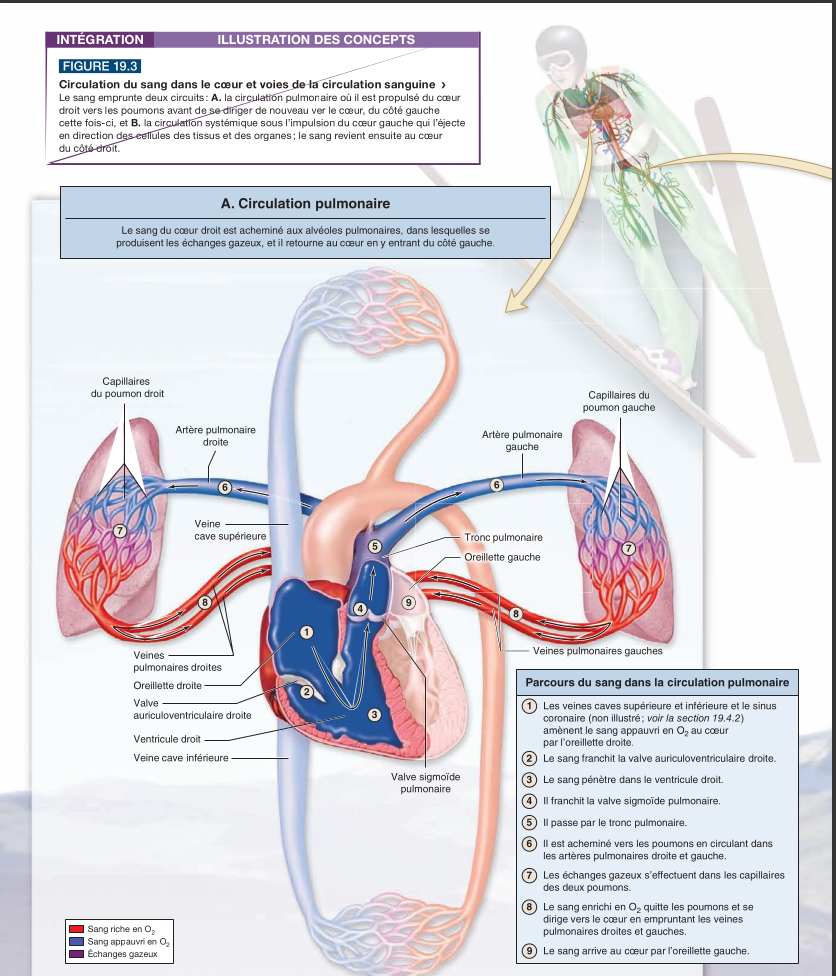
|
|
|
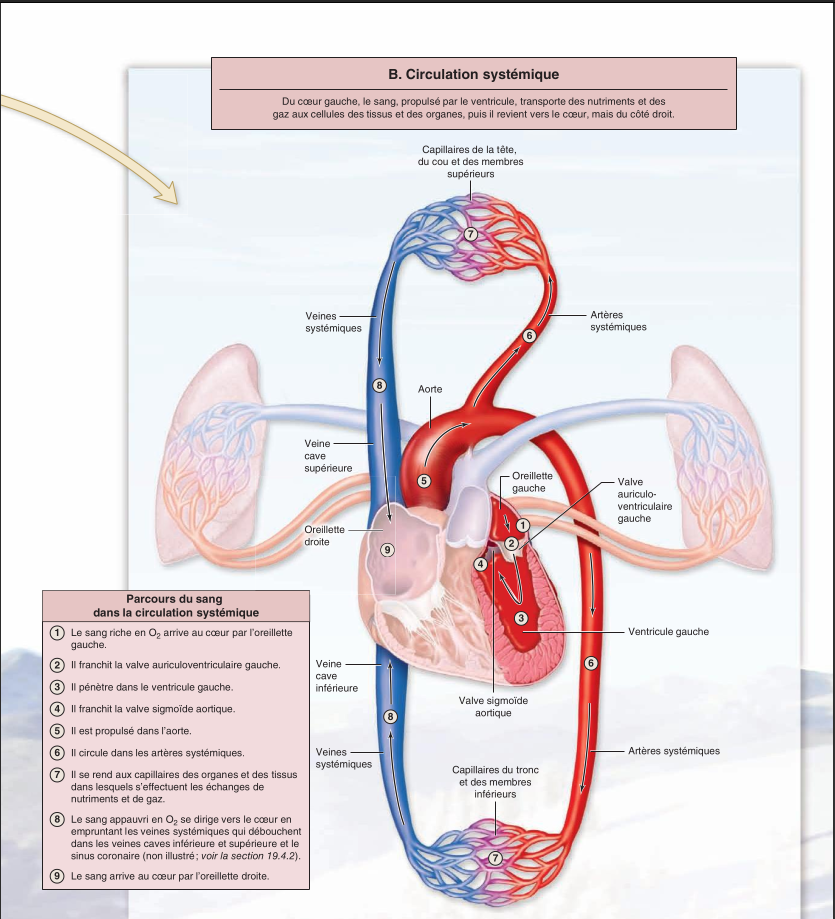
Du point de vue structurel, le système respiratoire se compose des voies respiratoires supé- rieures et des voies respiratoires inférieures. Du point de vue fonctionnel, le système respiratoire se compose de la zone de conduction et de la zone respiratoire.
Michael P. McKinley, Valerie Dean O'Loughlin et Theresa Stouter Bidle, Anatomie et physiologie, une approche intégrée, éd. McGraw-Hill/Chenelière éducation, 2014.
Les voies respiratoires supérieures se composent du nez, des fosses nasales et du pharynx.
Le nez débouche dans les fosses nasales. Cet espace, formé par le nez et le crâne, se divise en trois régions : le vestibule, la zone olfactive et la zone respiratoire.
Les fosses nasales se divisent en trois sections : le vestibule, la région olfactive et la région respiratoire. Trois des fonctions principales des fosses nasales consistent à réchauffer, à nettoyer et à humidifer l’air qui entre dans les voies respiratoires.
Les quatre sinus paranasaux sont reliés aux fosses nasales par des conduits. Ce sont les sinus frontaux, ethmoïdaux, sphénoïdaux et maxillaires. Ils permettent d'humidifier et de réchauffer l'air inhalé, et ils procurent une caisse de résonance à la voix.
Communément appelé gorge, le pharynx est un passage allongé mesurant en moyenne 13 centimètres. Il s’étend derrière les fosses nasales, la cavité orale et le larynx. L’air traverse le pharynx sur toute sa longueur ; la nourriture traverse uniquement sa partie inférieure. Les parois latérales du pharynx se composent de muscles squelettiques qui participent à la flexibilité et à la capacité d’extension de cette voie de passage, deux qualités qui lui permettent d’assurer la déglutition et de pousser les aliments vers l’œsophage.
Michael P. McKinley, Valerie Dean O'Loughlin et Theresa Stouter Bidle, Anatomie et physiologie, une approche intégrée, éd. McGraw-Hill/Chenelière éducation, 2014.
Les voies respiratoires inférieures regroupent toutes les structures qui s’échelonnent du larynx jusqu’aux alvéoles.
Le larynx sert de voie de passage pour l'air ; il ferme l'accès à la trachée au cours de la déglutition et contribue à la production des sons et à l'accroissement de la pression dans la cavité abdominale ; il intervient également dans les réflexes de l'éternuement et de la toux.
La trachée est un cylindre fexible qui va du larynx aux bronches principales ; elle est soutenue par des anneaux cartilagineux incomplets (en forme de C) qui maintiennent la trachée en état d'ouverture.
L'arbre bronchique est un système hautement ramifié de conduction de l'air qui se déploie depuis les bronches principales gauche et droite jusqu'aux bronchioles terminales.
Dans toutes les ramifications de l'arbre bronchique, la circulation de l'air est régie par la contraction et la détente du tissu musculaire lisse (la bronchoconstriction et la bronchodilatation, respectivement), surtout dans les bronchioles.
La zone respiratoire regroupe les régions des voies respiratoires dont la paroi est assez mince pour permettre les échanges gazeux, soit les bronchioles respiratoires, les conduits alvéolaires et les alvéoles.
Michael P. McKinley, Valerie Dean O'Loughlin et Theresa Stouter Bidle, Anatomie et physiologie, une approche intégrée, éd. McGraw-Hill/Chenelière éducation, 2014.
Les poumons se trouvent dans la cavité thoracique, de part et d'autre du médiastin ; la cage thoracique les entoure et les protège.
Le poumon est une structure conique divisée en lobes. Chaque poumon comprend un hile par lequel passent les bronches, les vaisseaux pulmonaires, les vaisseaux lymphatiques et les nerfs. Le poumon gauche comprend deux lobes, et le droit en compte trois.
La circulation pulmonaire achemine le sang désoxygéné jusqu'aux alvéoles (surfaces d'échanges gazeux des poumons), où il est rechargé en oxygène ; la circulation bronchique achemine le sang oxygéné jusqu'aux bronches et aux bronchioles.
Le tissu musculaire lisse du larynx et des bronchioles est innervé par le système nerveux autonome.
Michael P. McKinley, Valerie Dean O'Loughlin et Theresa Stouter Bidle, Anatomie et physiologie, une approche intégrée, éd. McGraw-Hill/Chenelière éducation, 2014.
Le volume d’air contenu dans le nez, la bouche, pharynx, bronches et bronchioles représente l’espace mort (150 mL) qui ne contribue pas aux échanges gazeux.
La ventilation pulmonaire
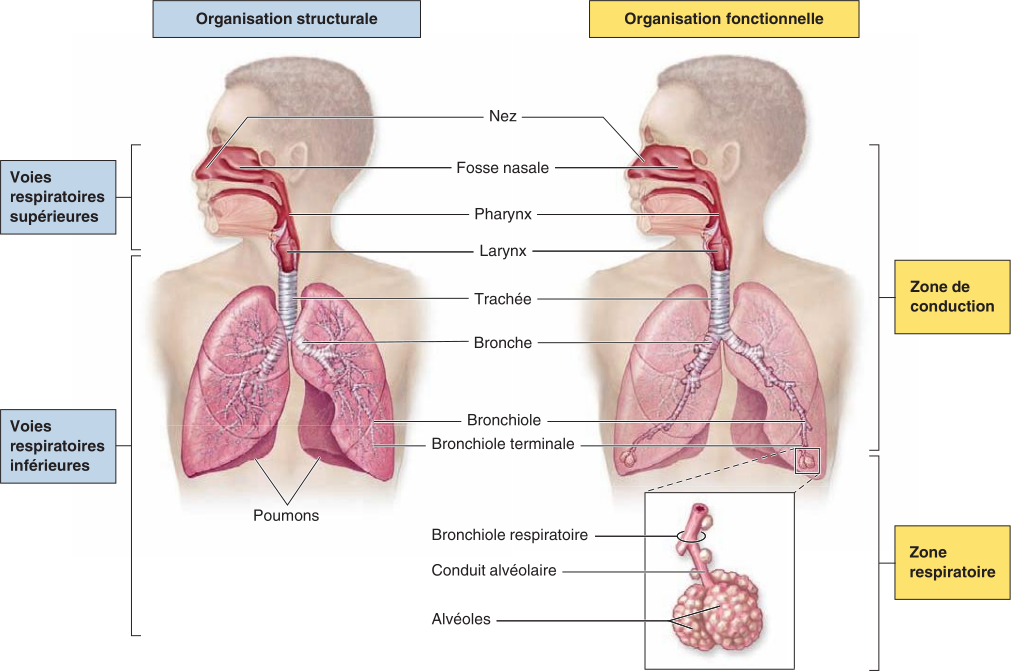
La ventilation pulmonaire se définit comme l'ensemble des interactions coordonnées des systèmes respiratoire, squelettique, musculaire et nerveux qui font entrer et sortir l'air des voies respiratoires.
À l'inspiration normale, le diaphragme et les muscles intercostaux externes se contractent pour augmenter le volume de la cavité thoracique et faire baisser la pression dans cet espace ; l'air se déplace dans le sens du gradient de pression descendant, de l'atmosphère jusque dans les alvéoles.
À l'expiration normale, les muscles qui se sont contractés pour l'inspiration normale se détendent pour réduire le volume de la cavité thoracique et augmenter la pression dans cet espace ; l'air se déplace dans le sens du gradient de pression descendant depuis les alvéoles jusque dans l'atmosphère.
La respiration forcée sollicite d'autres muscles qui se contractent pour augmenter davantage le volume thoracique et modifier la pression dans cette cavité.
La loi de Boyle-Mariotte stipule qu'à température constante, la pression (P) d'un gaz diminue quand le volume (V) du contenant augmente, et inversement. Elle peut ainsi s'exprimer de la manière suivante :
P1V1 = P2V2
où P1 et V1 représentent respectivement la pression et le volume à l'état initial, et P2 et V2 les représentent à l'état final.
La variation du volume thoracique au moment de l'inspiration et durant l'expiration induit un gradient de pression entre l'atmosphère et la cavité thoracique, et cet écart de pression détermine la circulation de l'air. Toute augmentation du volume de la cavité thoracique s'accompagne d'une baisse de la pression dans cet espace et provoque un affux de l'air dans les poumons (au moment de l'inspiration). À l'inverse, toute baisse du volume de la cavité thoracique s'accompagne d'une augmentation de la pression dans cet espace et provoque l'expulsion de l'air contenu dans les poumons (au moment de l'expiration).
La régulation nerveuse de la ventilation
La fréquence et l'amplitude (profondeur) de la respiration sont définies par des réflexes faisant intervenir les chimiorécepteurs, les propriocepteurs, les barorécepteurs et les récepteurs d'irritation. La respiration est également régie par les centres cérébraux supérieurs.
Les chimiorécepteurs mesurent les variations du pH induites par les fuctuations de la PCO2 du sang (il faut se rappeler que le CO2 peut mener à la production d'ions H+ acides). Lorsque les chimiorécepteurs détectent une augmentation de la concentration en ions H+ dans le sang, ils le signalent au centre respiratoire. Le centre respiratoire augmente alors la fréquence et la profondeur de la respiration dans le but de rétablir l'homéostasie.
Les chimiorécepteurs périphériques sont également stimulés par les variations de la PO2 du sang. D'une manière générale, les fluctuations de la PO2 du sang altèrent la fréquence respiratoire parce qu'elles rendent les chimiorécepteurs plus sensibles aux variations de la PCO2 du sang.
Le taux d'oxygène artériel dans le sang doit diminuer considérablement pour stimuler les chimiorécepteurs indépendamment de la PCO2 : la valeur de PO2 normale, qui est de 95 mm Hg, doit descendre à 60 mm Hg (ce qui est très faible). Ce faible seuil de stimulation des chimiorécepteurs entraîne parfois la mort de nageurs qui sont en état d'hyperventilation avant de s'immerger. L'hyperventilation abaisse en effet le taux sanguin de CO2 du nageur à un point tel que ses chimiorécepteurs ne sont plus stimulés (cela survient si la PCO2 tombe sous le seuil de 40 mm Hg). L'effort physique déployé durant la nage abaisse par ailleurs le taux sanguin de O2, mais pas suffisamment pour que les chimiorécepteurs soient stimulés. La baisse de PO2 dans les artères du cerveau du nageur peut alors mener à une perte de connaissance de ce dernier, entraînant ainsi des risques de noyade, avant que la PCO2 atteigne le niveau requis pour stimuler les chimiorécepteurs.
La PCO2 représente le stimulus le plus important dans l'évolution de la fréquence et de la profondeur de la ventilation pulmonaire. Le centre respiratoire est très sensible aux variations des niveaux de dioxyde de carbone : une augmentation minime de la PCO2 (de l'ordre de 5 mm Hg) peut multiplier par deux la fréquence respiratoire.
Les centres cérébraux supérieurs, soit l'hypothalamus, le système limbique et le cortex cérébral, peuvent également influer sur la fréquence respiratoire. L'hypothalamus (centre de contrôle important) augmente la fréquence respiratoire quand le corps a chaud, et la diminue quand il a froid. Le système limbique, participant aux émotions, altère la fréquence respiratoire en réaction aux émotions ou à des souvenirs chargés d'émotion. Le lobe frontal du cortex cérébral régit les modifications volontaires de la respiration pour les besoins de différentes activités, par exemple pour parler, chanter, retenir son souffle, appliquer la manœuvre de Valsalva, etc.
Les noyaux qui composent le centre respiratoire régissent la respiration normale en acheminant leurs impulsions nerveuses régulières par l'intermédiaire des nerfs phréniques et intercostaux. Le centre respiratoire modifie la fréquence et la profondeur de la ventilation pulmonaire en fonction de différents stimulus sensoriels qui lui parviennent. Le cortex cérébral assure quant à lui la régulation consciente de la respiration en stimulant directement les neurones qui se rendent jusqu'aux muscles squelettiques de la respiration. Cette diversité des influx nerveux régissant la respiration fait d'elle une activité à la fois consciente et non consciente (activité réflexe).
Michael P. McKinley, Valerie Dean O'Loughlin et Theresa Stouter Bidle, Anatomie et physiologie, une approche intégrée, éd. McGraw-Hill/Chenelière éducation, 2014.
La ventilation pulmonaire est la quantité d'air qui passe de l'atmosphère aux alvéoles en une minute. La ventilation alvéolaire correspond à la quantité d'air qui arrive aux alvéoles et qui peut participer aux échanges gazeux en une minute ; elle est inférieure à la ventilation pulmonaire, car une partie de l'air inspiré reste dans l'espace mort anatomique.
(Volume courant - espace mort anatomique) x fréquence respiratoire = ventilation alvéolaire
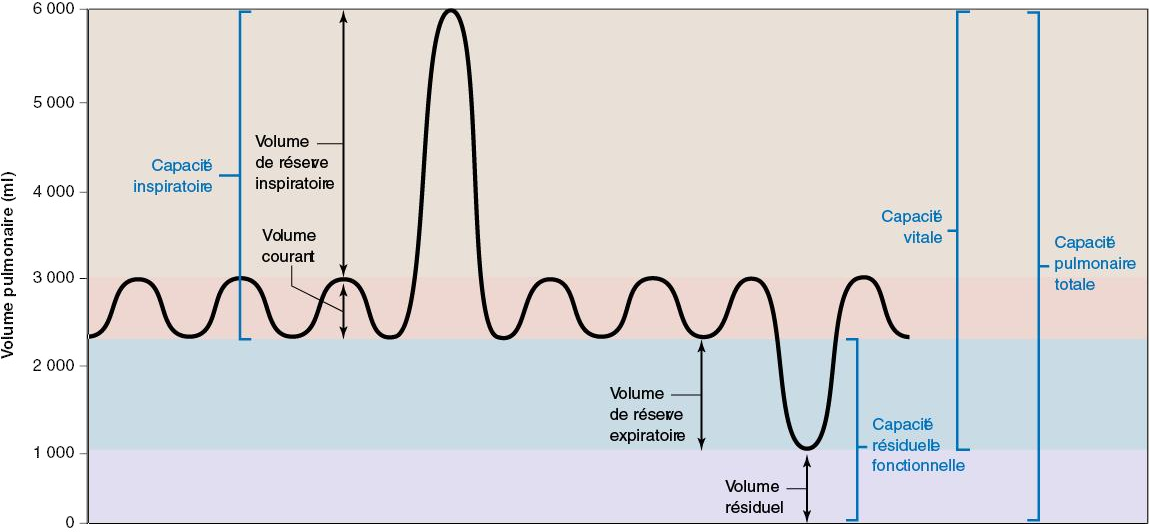
Volumes
Volume Définition Valeurs normales (homme) Valeurs normales (femme) Volume courant (VC) Quantité d'air inspiré ou expulsé des poumons au cours d'une respiration normale
500ml
500ml
Volume de réserve inspiratoire (VRI) Quantité d'air qui entre dans les poumons au cours d'une inspiration forcée, au terme d'une inspiration normale ; compliance pulmonaire mesurée par le VRI
3100ml
1900ml
Volume de réserve expiratoire (VRE) Quantité d'air expulsé des poumons au cours d'une expiration forcée, au terme d'une expiration normale ; élasticité des poumons et de la paroi thoracique mesurée par le VRE
1200ml
700ml
Volume résiduel (VR) Quantité d'air restant dans les poumons au terme d'une expiration forcée
1200ml
1100ml
Capacités
Capacité Formule Définition Valeurs normales (homme) Valeurs normales (femme) Capacité inspiratoire (CI) VC + VRI
Capacité totale d'inspiration
3600ml
2400ml
Capacité résiduelle fonctionnelle (CRF) VRE + VR
Quantité d'air restant normalement (résiduelle) dans les poumons à la n d'une expiration normale
2400ml
1800ml
Capacité vitale (CV) VC + VRI + VRE
Mesure de la puissance respiratoire
4800ml
3100ml
Capacité pulmonaire totale (CPT) VC + VRI + VRE + VR
Quantité totale d'air que les poumons peuvent contenir
6000ml
4200ml
Le volume expiratoire maximal (VEM) est le pourcentage de la capacité vitale qui peut être expulsé en un temps donné. La ventilation maximale minute (VMM) est la quantité maximale d'air qui peut être inspiré puis expulsé des poumons en une minute.
les échanges gazeux alvéolaires et systémiques
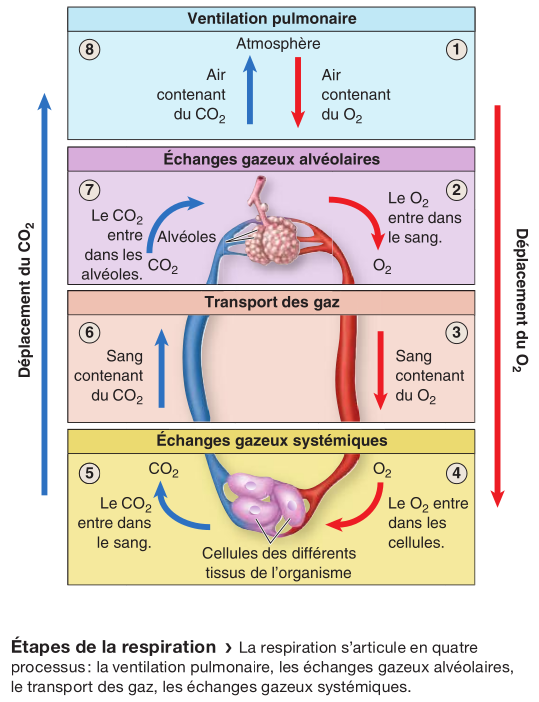
D'autres processus interviennent dans la respiration, notamment les échanges de gaz respiratoires entre les alvéoles et le sang (échanges gazeux alvéolaires), et entre le sang et les cellules systémiques (échanges gazeux systémiques).
La pression partielle de chaque gaz, ou sa contribution dans la pression totale, est égale à la pression totale exercée par le mélange gazeux, multipliée par le pourcentage qu’un gaz donné représente dans l’ensemble.
Pression totale × % d’un gaz dans le mélange = Pression partielle de ce gaz
Par conséquent, la pression partielle de chacun des gaz atmosphériques s’obtient à partir de la pression totale (qui s’établit pour l’atmosphère au niveau de la mer à 760 mm Hg) et du pourcentage de chacun des gaz les plus courants : l’azote (78,6 %), l’oxygène (20,9 %), le dioxyde de carbone (0,04 %) et la vapeur d’eau (0,46 %).
La loi de Dalton formalise la relation entre les pressions partielles et la pression totale : elle stipule que la pression totale d’un mélange gazeux est égale à la somme des pressions partielles de tous les gaz qui le constituent.
Quand la pression partielle d’un gaz est plus grande dans une région du système respiratoire que dans une autre, cet écart génère un gradient de pression partielle. Dans ce cas, le gaz se déplace de la région dans laquelle sa pression partielle est la plus élevée vers la région dans laquelle elle est la plus faible ; il conti- nue ainsi de se déplacer jusqu’à ce que les pressions partielles des deux régions soient devenues égales. Les échanges de gaz respiratoires se font ainsi dans le sens des gradients de pression partielle descendants, et ce, tant dans les échanges gazeux alvéolaires que dans les échanges gazeux systémiques.
Les gradients de pression partielle et le coefficient de solubilité des gaz déterminent en grande partie la solubilité des gaz dans un liquide.
D’autres principes chimiques régissent les échanges gazeux entre l’air, qui est un gaz, et le sang, qui est un liquide. La loi de Henry stipule les principes suivants : à une température donnée, la solubilité d’un gaz dans un liquide (c’est-à-dire la quantité de gaz qui peut entrer dans ce liquide ou en sortir) dépend de : 1) la pression partielle du gaz dans l’air ; et 2) le coefficient de solubi- lité du gaz dans le liquide.
La pression partielle du gaz constitue le moteur qui le fait entrer dans le liquide. Il faut se rappeler que la pression partielle dépend de la pression totale et du pourcentage que le gaz repré- sente dans le mélange gazeux en question ; si l’une ou l’autre de ces deux variables change, la quantité de gaz qui entre dans le liquide change également.
Le coefficient de solubilité est le volume de gaz qui se dissout dans un volume prédéterminé de liquide à une température et à une pression données. Cette constante dépend des interactions entre les molécules du gaz et celles du liquide. Plus ces interactions moléculaires sont nombreuses à une pression partielle donnée, plus la quantité de gaz qui se dissout dans le liquide est importante.
Tous les gaz ne sont pas également solubles dans l’eau. Ainsi, l’oxygène est très peu soluble dans l’eau (coefficient de solubilité = 0,024) ; le dioxyde de carbone est environ 24 fois plus soluble que l’oxygène (coefficient de solubilité = 0,57). Des trois gaz majeurs de l’atmosphère, l’azote est le moins soluble, soit presque deux fois moins que l’oxygène. Par ordre décrois- sant de solubilité, ces trois gaz se classent donc de la manière suivante :
CO2 > O2 > N2
La quantité de gaz qui peut se dissoudre dans un liquide dépendant à la fois de la pression partielle de ce gaz et de son coefficient de solubilité, les gaz les moins solubles doivent être soumis à des gradients de pression plus importants pour entrer dans le liquide.
Au niveau de la mer, à l'intérieur des alvéoles, la pression partielle de l'oxygène (PO2) est de 104 mm Hg et la pression partielle du dioxyde de carbone (PCO2) est de 40 mm Hg. Dans les cellules des tissus systémiques, la PO2 est de 40 mm Hg et la PCO2 est de 45 mm Hg.
Les échanges gazeux alvéolaires se définissent par le déplacement de l'oxygène et du dioxyde de carbone dans le sens de leur gradient de pression partielle descendant respectif.
L'efficacité des échanges gazeux alvéolaires est déterminée par la surface de membrane respiratoire disponible pour les échanges gazeux, par l'épaisseur de cette membrane et par le couplage ventilation-perfusion.
Les échanges gazeux systémiques se définissent par le déplacement des gaz respiratoires entre les capillaires systémiques et les cellules qui composent les tissus systémiques. Ils s'effectuent dans le sens d'un gradient de pression partielle descendant (de la pression la plus élevée vers la moins élevée).
Les échanges gazeux alvéolaires se produisent entre les alvéoles et les capillaires pulmonaires. À cet endroit, la PO2 du sang augmente et passe de 40 à 104 mm Hg. La PCO2 passe quant à elle de 45 à 40 mm Hg. Les échanges gazeux systémiques se produisent entre les cellules des tissus systémiques et les capillaires systémiques. À cet endroit, la PO2 du sang diminue et passe de 95 à 40 mm Hg. La PCO2 passe quant à elle de 40 à 45 mm Hg.
Le transport des gaz
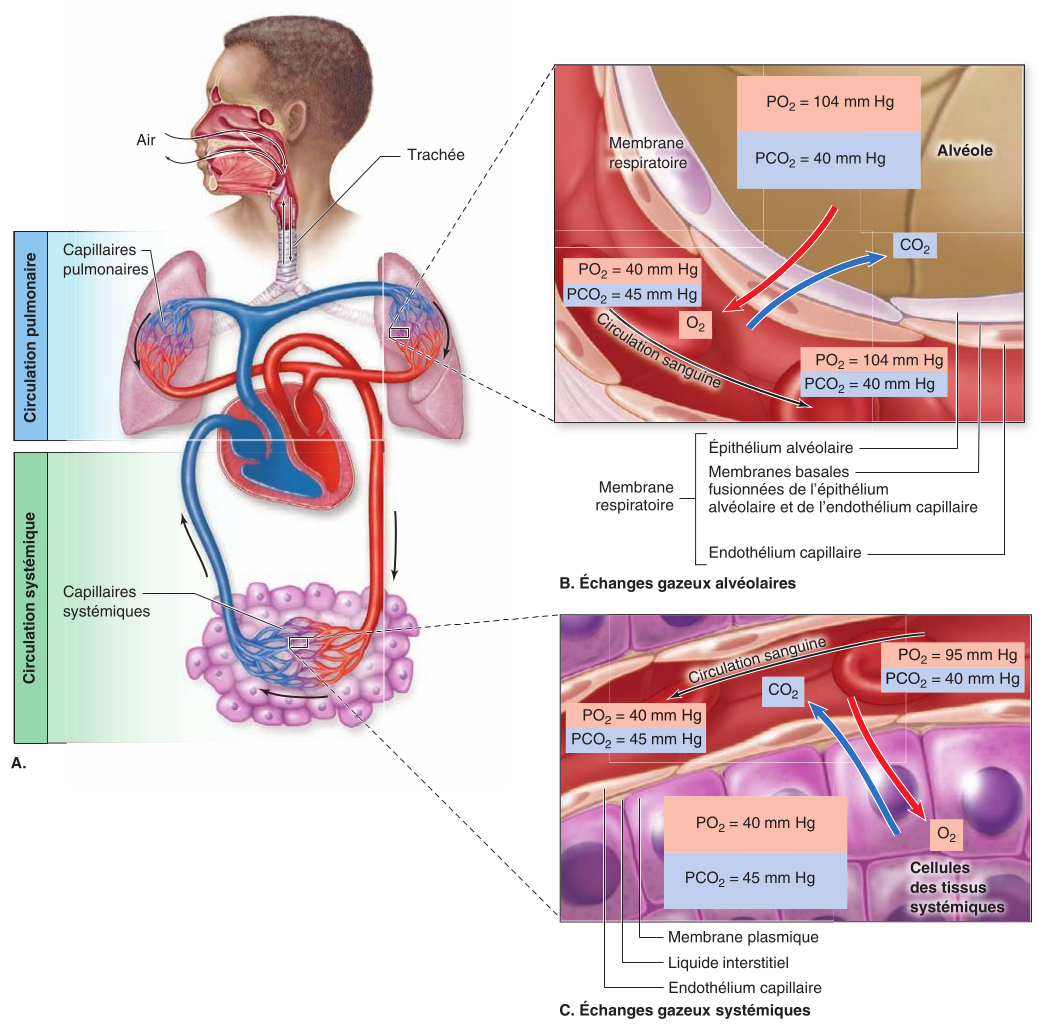
Le transport des gaz correspond aux déplacements des gaz respiratoires entre les poumons et les cellules systémiques par le débit sanguin.
Pour être acheminé par le sang, l'oxygène se lie au fer de l'hémoglobine (plus de 98 % de la quantité totale d'oxygène) et se dissout dans le plasma (moins de 2 %).
Le dioxyde de carbone est principalement acheminé sous forme d'ions HCO3 – (70 % de la quantité totale) ; il peut aussi être lié à la globine de l'hémoglobine (23 %) ou dissous dans le plasma (7 %).
À l'intérieur de l'érythrocyte, le CO2 réagit avec le H2O pour former le H2CO3, qui s'ionise en HCO3− et en H+.
L'hémoglobine transporte l'oxygène, le dioxyde de carbone et les ions H+.
La PO2 constitue le facteur le plus déterminant du transport de l'oxygène par l'hémoglobine. La saturation de l'hémoglobine en oxygène augmente avec la PO2.
D'autres variables stimulent la libération de l'oxygène (donc inhibent la liaison de l'oxygène à l'hémoglobine), notamment l'élévation de la température, la baisse du pH, l'augmentation du nombre de molécules de 2,3-DPG et l'accroissement du taux de dioxyde de carbone.
La fréquence respiratoire et l'homéostasie
La fréquence respiratoire détermine en partie la PO2, la PCO2 et le pH du sang.
La fréquence respiratoire influe également sur le retour veineux du sang et de la lymphe.
L'hyperventilation fait baisser la PCO2 du sang et peut ainsi augmenter le pH.
L'hypoventilation fait baisser la PO2 du sang et augmenter la PCO2, ce qui peut mener à une diminution du pH.
La contraction et le relâchement des muscles squelettiques de la respiration provoquent des variations de pression régulières sur les vaisseaux sanguins et lymphatiques, ce qui contribue à la circulation du sang et de la lymphe.
L'approfondissement de la respiration attribuable à l'effort physique s'appelle l'hyperpnée.
Michael P. McKinley, Valerie Dean O'Loughlin et Theresa Stouter Bidle, Anatomie et physiologie, une approche intégrée, éd. McGraw-Hill/Chenelière éducation, 2014.
Les humains et autres mammifères ont une réponse à l'immersion consistant en un ensemble de réflexes qui sont activés lorsque notre visage est refroidi (comme par l'eau lors d'une plongée) ou si nous retenons notre souffle. Le réflexe d'immersion est un astucieux mécanisme physiologique permettant à l'organisme de tolérer un faible niveau d'oxygène. Ceci est en partie réalisé par un rythme cardiaque plus faible et en partie par une constriction des vaisseaux sanguins périphériques dans les bras et les jambes pour diriger le sang vers les organes internes vitaux comme le cœur et le cerveau qui ont le plus besoin d'oxygène. Les changements dans le corps se produisent relativement rapidement, dans les 30 secondes.
Stig Severinsen, Breatheology : The Art of Conscious Breathing, éd. Glaucus Pub Ltd Inc, 2009.
C'est principalement le système cardiovasculaire qui permet des plongées incroyables. De nombreuses réponses de plongée humaine ont également été observées chez les mammifères marins : le sang qui irrigue la périphérie du corps est rappelé au cœur du torse, et la fréquence cardiaque est réduite, assurant une plus grande économie d'utilisation de l'oxygène. Le phoque par exemple, qui peut plonger à environ 300 mètres, ralentit son pouls de 120 battements par minute à 20.
Plusieurs études d'Umberto Pelizzari ont révélé que dans une apnée stationnaire dans l'eau, sa fréquence cardiaque ralentit progressivement à 30 battements par minute. Le phénomène est encore plus évident lors d'apnée en profondeur.
Umberto Pelizzari et Stefano Tovaglieri, Manual of freediving, éd. Idelson-Gnocchi Ltd., 2004.
Après plusieurs plongées, la rate se contracte et libère une grande quantité de globules rouges dans le système circulatoire. La contraction de la rate se produit beaucoup plus lentement que les autres réflexes de plongée. La libération de plus de globules rouges permet de stocker plus d'oxygène dans le sang. Enfin, la quantité supplémentaire de cellules sanguines permet au corps de retrouver son équilibre normal plus rapidement après une apnée prolongée. Pour parler de manière populaire, la rate agit comme une sorte de "turbo" - pendant et après une longue plongée.
Stig Severinsen, Breatheology : The Art of Conscious Breathing, éd. Glaucus Pub Ltd Inc, 2009.
Le sang est rappelé des zones périphériques du corps, où il n'y a pas d'organes vitaux, et est poussé dans les poumons, où il occupe l'espace libre laissé par la réduction du volume d'air due à l'augmentation de la pression. Le sang est un liquide et donc incompressible ; cela explique comment nous pouvons nous adapter à la pression sans imploser.
Mais le transfert sanguin n'est pas seulement un phénomène passif qui s'oppose à la pression hydrostatique ; c'est aussi un phénomène actif qui permet une exploitation plus rationnelle de l'oxygène, le réservant pour les organes critiques comme le cerveau et le cœur, à la perte d'organes et de tissus périphériques.
Umberto Pelizzari et Stefano Tovaglieri, Manual of freediving, éd. Idelson-Gnocchi Ltd., 2004.